
Angiosperm: Reproductive structures
General features of reproductive structures:
- In Angiosperms, there is a wide range of morphology and structure of the reproductive organs of the plant.
- Flowers being the reproductive tissues of the plant comprises of both the male and female reproductive organs in it.
- They may be found at the termination of short lateral branches or the main axis or both.
- Flowers can be found either individually as in daffodils or in clusters termed as inflorescence as in sunflower.
- The floral parts of the angiospermic plants give fruits.
- A complete flower comprises of the four organs that are attached to the floral stalk via the receptacle.
- The four organs are sepals, petals, stamens and carpels which is located above the base of the receptacle.
- In case of dicots, the organs are usually grouped in the multiples of four and five, whereas, in case of monocots, the organs are grouped in multiples of three.
- The sepals are the outermost layer and are usually green in colour.
- The sepals encloses the flower bud and are collectively termed as calyx.
- The next layer of floral appendages inside the calyx are petals.
- Petals are generally bright in color and are collectively termed as the corolla.
- Jointly, the calyx and corolla form the perianth.
- Even if the sepals and petals protect the flower buds and attract the pollinators, they do not take part directly in sexual reproduction.
- Thus, they are termed as accessory parts.
- In case if the color and the appearance of sepals and petals are identical, then the perianth is said to be composed of tepals. For example: Easter Lily (Lilium longiflorum).
- The stamens are the spore producing structures (microsporophylls) and are located interior to the corolla.
- The stamens are collectively termed as androecium.
- The stamens comprise of a slender stalk (the filament) in most angiosperms, which contains the anther (and pollen sacs) inside which the pollen is produced.
- At the base of the stamens, tiny secretory structures called nectaries are usually found which supply food rewards for pollinators.
- The nectaries unify into a nectary or staminal disk in certain cases.
- In certain cases, as a whorl of stamens is reduced into a nectiferous disk, the staminal disc develops, and in some cases, the staminal disc is simply produced from the receptacle’s nectary-producing tissue.
- Megasporophylls are termed as carpels.
- Carpels enclose one or more ovules, each with an egg.
- The ovule matures into a seed after fertilization, and the carpel grows into a fruit.
- Carpels, and hence fruits, are special to angiosperms.
- A flower is termed to be complete when it possess all four organs, whereas, it is termed as incomplete when any one of it is missing.
- Both stamens and carpels are present in a bisexual (or “perfect”) flower.
- A unisexual (or “imperfect”) flower either lacks stamens and is termed as carpellate or lacks carpels and is termed as staminate.
- The term monoecious is given for the species where both the carpellate and staminate flowers are on the same plant.
- The term dioecious is given for the species where staminate flowers are on one plant and carpellate are present on the other.
- Floral organs are usually open or fused.
- The fusion of similar organs is termed as connation eg. the fused petals as in morning glory.
- The fusion of different organs is termed as adnation eg. the fusion of stamens to petals in the mint family.
- The basic floral pattern comprises of the alternating whorls of the organs located concentrically from outside to inside as sepals, petals, stamens, and carpels, etc.
What are the reproductive structure of angiosperm?
1. Receptacle:
- The axis (stem) to which the floral organs are connected is termed as receptacle.
- The attachment of the floral organs are either in alternating successive whorls, found in majority of angiosperms or in a low continuous spiral form, as is common among primitive angiosperms.
- The stalk of a flower or of an inflorescence is termed as the peduncle.
- The peduncle is the internode between the receptacle and the bract when a flower is borne individually, where bract is the last leaf, usually modified and smaller in comparison to other leaves.
- In case where the flower are borne in an inflorescence, the internode between the receptacle of each flower and the bracteole is termed as a pedicel.
- Hence, in case of inflorescence, the pedicel is equivalent to peduncle whereas, bracteole is equivalent to bract.
- Usually the bract that subtends an inflorescence is bright in color like in the case of poinsettia (Euphorbia pulccherima) or provides the protection by woody boat shaped bracts in the cases of palms.
- In some angiosperms, as in strawberries the edible fleshy part is the receptacle.
- This, when eaten by birds and mammals helps in seed dispersal.
- The fleshy portion of the edible fruit forms from the receptacle and peduncle in cacti (e.g., prickly pear), and several internodes below which develop up and cover the carpels; this is why there are axillary buds on the fruit surface in cacti (areoles) with spines.
2. Calyx:
- Owing to their usually green colour, the sepals (collectively called the calyx) most resemble leaves.
- Sepals remain separate (aposepalous or polysepalous) or slightly fused (synsepalous) from their base and along much of their length, forming a tube with terminal lobes or teeth.
- The number of calyx lobes equals the number of sepals that are fused (connate).
- The sepals enclose the unopened bud of the flower and protect it.
- In comparison to the more short-lived petals and stamens, the calyx is usually persistent and visible as the fruit matures (e.g., persimmon, Diospyros virginiana).
- When true petals are absent, sepals can be brightly colored and act as petals, as in the virgin’s bower (Clematis) and the Bougainvillea.
- In this case, petaloid sepals vary from tepals because the first group of stamens is on the same radii as the sepals, indicating the absence of petals, which in the next floral whorl will usually be located on alternating radii.
3. Corolla:
- The petals constituting the corolla are usually brightly coloured or white and lure insects and birds for pollination.
- Typically, the number of petals is identical to the number of sepals.
- The petals describe floral symmetry.
- The flower has radial symmetry when the petals of the corolla are of the same shape and when they are equidistant from each other and the flower is named regular or actinomorphic.
- In regular flowers, the flower will be split into two identical halves by any line drawn across the middle.
- The flower has bilateral symmetry and is called irregular or zygomorphic if at least one petal of the corolla is different. Eg. violets.
- For all or part of their length, the corolla petals may be separate, or apopetalous, or marginally fused (fusion of similar floral parts is called connation), or sympetalous.
- They form a tubular corolla with terminal lobes when joined.
- In regular flowers e.g., blueberries or irregular flowers, eg. sage, a tubular corolla may be present.
- Stamens are generally associated with a tubular corolla.
- A marginally fused calyx is termed as synsepalous.
- A marginally fused corolla is termed as synpetalous.
- Synsepalous and synpetalous along with stamens fuse to form a cup like floral tube called a hypanthium that surrounds the carpel. For ex. Cherries.
- Fusion and reduction of flower parts are more frequent and have taken place in several unrelated lineages.
- Several wind-pollinated angiosperms lack petals, nor have floral parts modified as petals; the amaranth family and the birch family are examples of wind-pollinated plants.
- Petals also hold nectaries that secrete compounds containing sugar, and petals often develop fragrances to attract pollinators; petals derive the scent of a rose.
- Petals also produce an extension of the tubular corolla containing nectar, called a spur.
- This may contain one petal, as in the larkspur, or all the petals, as in columbine, both being the members of the family Ranunculaceae.
4. Androecium:
- Stamens (microsporophylls) are pollen producing structures present in terminal saclike structures (microsporangia) termed as anthers.
- Sometimes the number of stamens comprised by the androecium is the same as the number of petals, but sometimes the stamens are more or less numerous than the petals.
- In a young stamen, there are usually two pairs of spore-containing sacs (microsporangia); the distinction between the adjacent microsporangia of a pair breaks down during maturation so that there are only two pollen-containing sacs (one in each anther lobe) at the time the pollen is released by the stamen.
- The less modified stamens, with the paired microsporangia located near the margins, are identical to leaves; an example is found in the magnolia family.
- The blade becomes modified into a slender stalk, the filament, with the microsporangia at or near the apex of the filament in more derived stamens.
- Generally, the filaments are attached with the corolla, but either isolate with the anthers, as in primroses (Primula; Primulaceae), or merged with each other to form a staminal tube enclosing the gynoecium, as in the mallow family.
- The staminal tube is fused with the lower half of the corolla tube in the thistle (Cirsium; Asteraceae) and in other members of the sunflower family.
- In stamen modification, there are many patterns.
- One or more of the stamens in many angiosperms are modified and lack functional anthers.
- The filament is extended in the most common modification to form a petal-like blade called a staminode.
- Apparent petals are of staminodial origin in some angiosperm families as seen in many members of Caryophyllaceae.
- Wild roses have only five petals and several stamens, but for the many apparent petals (but actually staminodes) and few usable stamens, cultivated roses have been selected.
- Stamens have been transformed into sterile nectaries involved in pollination in other situations.
- If flowers have a large number of stamens, then, as in the myrtle family, the stamens frequently occur in groups or clusters.
5. Gynoecium:
- Gynoecium is made up of carpels.
- Carpels are spirally arranged in more basal families (e.g., Magnoliaceae), and in more advanced families they appear to be arranged in a single whorl.
- The number of carpels ranges from one (e.g. Fabaceae family) to several (e.g. Raspberries).
- The ovary is at the base of a carpel, inside which one or more multicellular structures called ovules develop, each containing an egg.
- The pollen is received by the upper portion of the carpel is termed as the stigma.
- The ovary and the stigma are often connected by a slender stalk called the style.
- The carpels may be free (apocarpous) or fused (syncarpous), with the walls and cavities (locules) of the individual carpels still present.
- As in the wood sorrel (Oxalis), syncarpy may include only the ovaries, leaving the styles and stigmas free, or it may include both the ovaries and styles, keeping only the stigmas free, as in the waterleaf.
- The number of carpels in the syncarpous (or compound) ovary is generally identical to the number of locules.
- The location of the gynoecium on the floral axis with regard to the petals, sepals, and stamens also characterizes the flower.
- The perianth and stamens are connected to the receptacle below the gynoecium in hypogynous flowers; the ovary is superior to these organs, and the remaining floral organs originate from below the carpel’s point of origin.
- A hypanthium (a floral tube developed from the fusion of the stamens, petals, and sepals) is attached to the receptacle below the gynoecium in periginous flowers and surrounds the ovary; the ovary is superior, and the free parts of the petals, sepals, and stamens are attached to the hypanthium surface.
- The hypanthium is fused to the gynoecium in epigynous flowers, and the free parts of the sepals, petals, and stamens tend to be attached to the top of the gynoecium, as in the apple (Malus; Rosaceae); the ovary is inferior, and from the top of the ovary the petals, sepals, and stamens seem to emerge.
6. Fruit:
- Fertilization of an egg by a compatible pollen grain inside a carpel results in the development of seeds within the carpel.
- A ripened ovary (or compound ovary) and some other structure, usually the hypanthium, which ripens and forms a unit with it s termed as fruit .
- The formation of fruit without the fertilization of an egg and subsequent seed production is termed as parthenocarpy.
- As a vegetable is produced only from vegetative (non-reproductive) organs, this specifically distinguishes a fruit from a vegetable.
- Some examples of fruits are tomatoes, squashes, eggplants as they are derived from floral parts.
- From one single carpel or from a compound ovary, simple fruits grow.
- The aggregate fruits comprise of several single apocarpous gynoecium carpels. ex. Raspberries.
- In multiple fruits, gynoecia of more than one flower are found and represent a complete inflorescence as such fig and pineapple.
- In the development of the mature fruit, accessory fruits incorporate other flower parts; for instance, the hypanthium is used to form the pear (Pyrus; Rosaceae), and the receptacle becomes part of the prickly pear.
- Fruit shape, texture, and composition are variable (notably in simple fruits), but most of them fall into a few categories.
- There are three layers of the fruit wall or pericarp i.e. endocarp being the inner layer; the mesocarp being the middle layer; and the exocarp being the outer layer.
- These layers can be either fleshy or dry (sclerified) or either of the two variations, however they are either classified as one or the other.
- Berries, drupes, and pomes are the three primary types of fleshy fruits.
- Berries are simple fruits with several seeds, consisting of one carpel or a syncarpous ovary.
- Throughout, they are fleshy, but the texture of the exocarp varies: a smooth thin exocarp, like in tomatoes (a berry); a leathery exocarp, like in oranges (a hesperidium); and a very stiff exocarp, like in pumpkins.
- Typically, only one seed per carpel or locule is found in drupes, or stone fruits.
- Drupes are fleshy fruits consisting of an inner stony or woody endocarp adhering to the seed as in peaches and cherries.
- For each aggregate fruit unit of this sort, the word druplet is used. eg. raspberries.
- Pomes are fleshy fruits belonging to the rose family (Rosaceae) where the adnate hypanthium is fleshy.
- Simple dry fruits can be either dehiscent or indehiscent.
- If the pericarp splits open at maturity and releases the seeds, they are dehiscent or indehiscent while the pericarp stays intact when the fruit is shed from the plant.
- Follicles, legumes, and capsules are the three main forms of dehiscent fruits.
- From either single carpels or compound ovaries, indehiscent fruits are produced.
- The achene, the samara, and the caryopsis are single carpel forms.
- Nuts and schizocarps include forms derived from a compound ovary.
- An achene is a fruit in which the single seed in the cavity lies free, only connected by a single point.
- For instance, the strawberry is indeed an aggregate fruit, and each ‘seed’ is an achene.
- In the tree of heaven (Ailanthus altissima; Simaroubaceae) and ash, the samara that is a winged achene is found.
- The seed adheres to the fruit wall in the caryopsis, or grain.
- Among the cereal grasses, such as corn, the caryopsis is found.
- Nuts have a stony pericarp and as in oak acorns (Quercus; Fagaceae) and hazelnuts, typically only a single seed matures in each carpel.
- Schizocarps are fruits that divide each carpel of a compound ovary into two or more components, each with a single seed.
- In the carrot family, schizocarps are found.
- In maples, winged schizocarps are found.
7. Seed:
- The mature ovules are seeds.
- For the seedling, they provide the developing embryo and the nutritive tissue.
- Seeds are surrounded by one or two integuments that grow into a typically hard seed coat.
- They are found in a carpel’s ovary and are thus protected from components and predators.
- The ovule is connected by a short stalk called the funiculus to the ovary wall before maturity.
- The area of connection to the ovary wall is termed as the placenta.
- The placental arrangement (placentation) in the angiosperm compound ovary is distinguished by the presence or absence of a central column in the ovary and the location of attachment.
- The placentae are positioned on a central column in axil placentation; partitions form chambers (locules) from the central column to the ovary wall that separate the placentae and attached ovaries from each other.
- Free-central placentation is similar to axile placentation, except the column is not attached to the ovary wall by partitions, so no locules are created.
- The ovules are connected to the base of the ovary in basal placentation, and the placentae are placed directly on the ovary wall or on its extensions in parietal placentation.
- Mature seeds are covered in integuments that could become stiff and stony, or that may have an outer fleshy sarcotesta with an inner stony sclerotesta, typically brightly colored.
- Seed coats might also be winged or variably ornamented with prickles or sclerified hairs.
- There may be an extra covering in certain seeds, the aril, which is an outgrowth of the funiculus.
- The tomato becomes slippery because of the aril.
8. Inflorescence:
- The clusters of flowers on a branch or system of branches is termed as inflorescence.
- On the basis of timing of their flowering and their arrangement on the axis, they are usually categorized.
- In case of indeterminate inflorescence, the youngest flowers that are last to be opened are placed at the top of the inflorescence in elongated axes, however, they are arranged in the center in case of truncated axes.
- At any distance from the main stem, branching and the associated flowers develop.
- There are diverse kinds of indeterminate inflorescences. They are racemes, panicles, spikes, catkins, corymbs, and heads.
- Racemes: A raceme is a type of inflorescence where a flower develops at the axil of each leaf through the elongated, unbranched axis.
- A short stalk called a pedicel terminates each flower.
- There is indeterminate growth of the main axis; thus the growth does not stop at the beginning of flowering.
- Spike: A spike is a raceme other than that the flowers are connected directly to the axis at the axil of each leaf instead of being attached to a pedicel.
- Cattail(Typha) is the example of spike.
- A spadix is considered the fleshy spike characteristic of the Araceae, and a spathe is referred to as the underlying bract.
- Catkin: A spike in which all the flowers are of only one sex, either staminate or carpellate, is termed as catkin (or ament).
- The catkin is normally pendulous and when the inflorescence as a whole is shed, the petals and sepals are reduced to assist in wind pollination. Ex:Oaks
- First the lower flowers open, and a corymb’s axis begins to produce flowers. Ex: hawthorn
- Corymbs: Corymbs are found in the hawthorn (Crataegus; Rosaceae). The flowers emerge from a common point and tend to be at about the same level if the axis is short or stunted.
- An umbel, is actually a flattened raceme as the internodes of the axis, or peduncle (the point of origin of the leaves and flower axes), are shortened so that the pedicels are of the identical length (eg. the carrot family).
- Head: A head is a raceme where the peduncle is flattened and the flowers are directly attached to it. Ex. Aster family.
- This leads to grouping of small flowers that are arranged so as they seem as a single flower.
- The ray (external) flowers have a well-developed zygomorphic corolla in several members of the Asteraceae (e.g., sunflowers), and the disk (internal) flowers have a small actinomorphic corolla.
- Normally, the inner disk flowers are complete flowers, and usually, the ray flowers are sterile.
- The main axis is branched in the compound indeterminate inflorescences such that the various inflorescences form off the main axis.
- A panicle is a branched raceme where the branches are racemes themselves. Eg: yuccas.
- The shift to flattened axes (corymbs and umbels) from elongated axes (racemes and panicles) results in inflorescences in which the flowers are placed close together. ex. Wild carrot.
- With compound spikes, catkins, corymbs, and heads, this organization is the same.
- The shift to flattened axes (corymbs and umbels) from elongated axes (racemes and panicles) results in inflorescences in which the flowers are placed close together.
- This close association facilitates successful pollination, and an inflorescence that appears to be a single flower is created by the intense condensation of the inflorescences, as in the head. Ex: sunflowers
- The youngest flowers are at the bottom of an elongated axis or on the outside of a truncated axis in the determinate (cymose) inflorescences. Ex. In the cymose umbel of onions.
- These inflorescences are determinate because the entire apical meristem generates a flower at the time of flowering; therefore the entire axis stops to develop.
- Each unit of the cyme includes dichasium, which consists of a central flower and two lateral flowers.
- The branching is mainly sympodial, and it may be compound inflorescence. Ex: catchfly.
- There is a one-sided cyme called a helicoid cyme in several monocotyledons.
- The cymose inflorescence if arranged at the nodes in pair, in the manner of false whorl is termed as verticillaster.
- Lastly, there are mixed inflorescences as for example, the cymose clusters arranged in a racemose way (eg. lilac) or other type of combinations.
Reproduction in angiosperms:
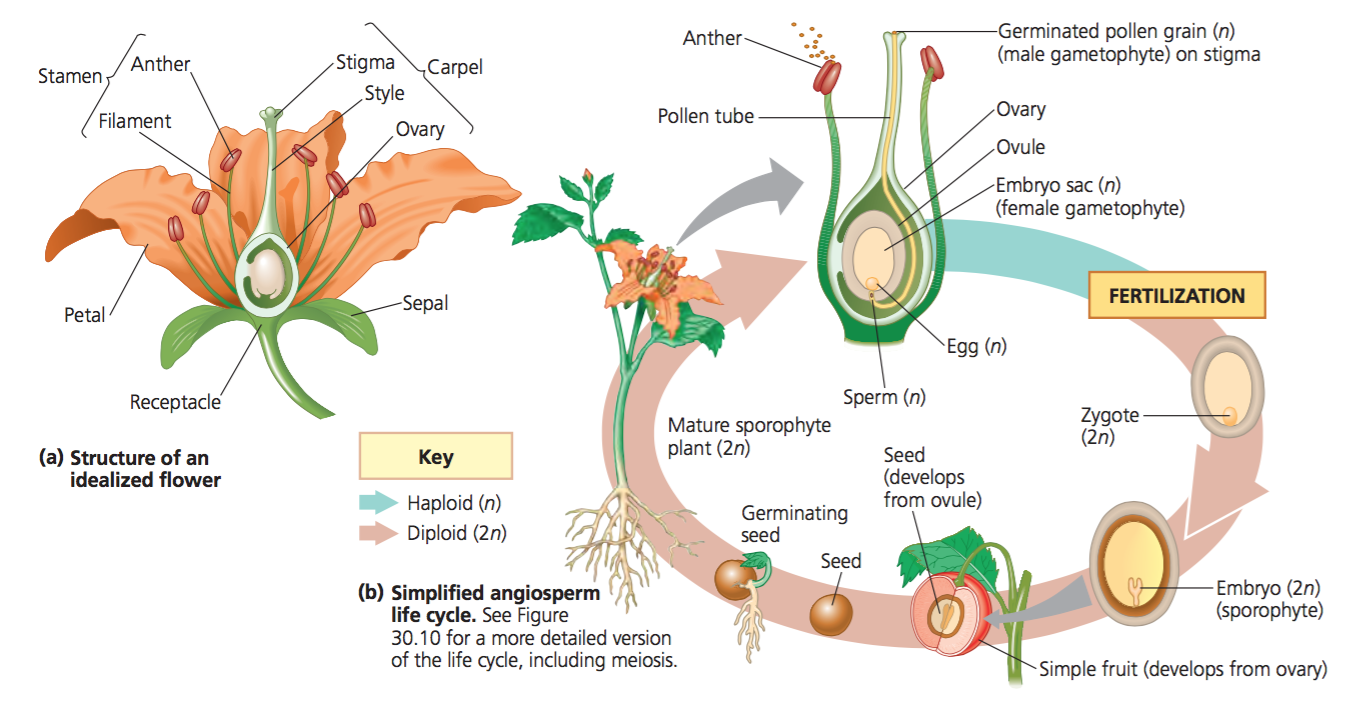
source: Apbiology
General features:
- The immense arrangement of the floral parts of the angiosperms is for the sexual reproduction.
- The life cycle of angiosperms comprises of sporophytic and gametophytic phase.
- The cells of sporophyte body is diploid (2n), and the sporophyte is the body of the plant that we observe.
- When preparing for the reproduction, the sporophyte undergoes meiotic cell division and gives rise to the gametophyte.
- Gametophyte are the reproductive cells that are haploid(n) in nature.
- Pollen grain is a two-celled stage microgametophyte that germinates into a pollen tube and gives haploid sperms via division.
- The embryo sac which is an eight-celled stage gives rise to the eggs.
- Being vascular plants, angiosperms have life cycle in which sporophyte phase is dominant and gametophyte is recessive.
- The sporophyte is green and photosynthetic whereas gametophyte relies on the sporophyte for the nourishment.
- The micro-gametophyte is reduced to 3-celled stage and the mega-gametophyte is of 8-celled stage in case of angiosperms.
- Pollination acts as a driver for the sexual reproduction by bringing these gametophytes in close association and hence facilitates for the fertilization.
- Pollination is referred as a process in which the pollens generated by the anthers is received by the stigma of the ovary.
- Fertilization takes place by the fusion of sperms and eggs in order to produce a zygote, which ultimately forms the embryo.
- The ovule develops into a seed after the fertilization and the ovary develops into a fruit.
a. Anthers:
- Four areas of tissue capable of developing spores are exposed by a transverse segment of the anther.
- These tissues are comprised of microsporocytes.
- Microsporocytes are diploid cells that are go through meiosis to form a tetrad of haploid microspores.
- The microspores become pollen grains and may separate eventually.
- The layer of cells below the dermis of the anther wall (the endothecium) develops thickness in the cell walls during pollen growth.
- The cell layer develops into a layer of nutritional cells immediately within the endothecium (the tapetum) that either secrete their contents into the region around the microsporocytes or lose their inner cell walls, separate from each other, and become amoeboid among the microsporocytes.
- The pollen grain develop a thick wall of two layers i.e. intine and exine.
- The intine is the inner layer that comprises of basically of cellulose and pectin.
- The exine is the outer layer that is comprised of sporopollenin.
- Sporopollenin is the highly decay resistant chemical.
- The exine posses one to many pores through which pollen grain germinates whereas the thick area of the exine is highly shaped.
- To form a two-celled microgametophyte, each microspore (pollen grain) divides mitotically.
- One cell is a tube cell (the cell that grows into a pollen tube), and the other is a generative cell, which, as a result of further mitotic division, will give rise to two sperm.
- Therefore, only three haploid cells, the tube cell and two sperm, form a mature microgametophyte.
- At the two-celled stage, most angiosperms shed pollen, however, in some advanced cases, it is shed at the mature three-celled stage.
- After the maturation of pollen grains, the anther wall breaks either longitudinally or by an apical pore.
b. Ovule:
- Ovule is a sac like structure that is enclosed by layers of cell.
- It is responsible for production of megaspores.
- In angiosperms, the nucellus is termed as the megasporangium.
- One or two integuments arise close to the base of the ovule primordium after the initiation of the carpel wall, expand in a rimlike manner, and enclose the nucellus, that leaves only a small opening called the micropyle at the top.
- The existence of two integuments in angiosperms is plesiomorphic (unspecialized) and one integument is apomorphic.
- The existence of two integuments in angiosperms is plesiomorphic (unspecialized) and one integument is apomorphic.
- Three of the four megaspores degenerate, and the one that remains enlarges.
- The resulting megagametophyte generates the female gametes.
- Free-nuclear mitotic divisions are involved in this development (called megagametogenesis).
- The cell wall remains intact until the megagametophyte, or embryo sac, is formed, while the nucleus divides.
- There are usually eight nuclei in the embryo sac.
- In gametophyte formation in gymnosperms, free-nuclear mitotic division is also found.
- To either end of the embryo sac, four nuclei migrate.
- Then, one nucleus from each group migrates to the embryo’s center; they become the polar nuclei.
- The two polar nuclei combine in the centre of the embryo sac to form a fusion nucleus.
- To form three antipodal cells, cell walls form around each of the chalazal nuclei.
- Enlargement of the embryo sac during development leads to the loss of much of the nucellus.
- In 70 percent of the angiosperms in which the life cycle has been charted, this series of megasporogenesis and megagametogenesis, called the Polygonum type, occurs.
- Differences present in the remaining 30 percent shows derivations from the Polygonum type of seed development.
Pollination:
- Efficient pollination involves the transition to a stigma of the same species of pollen from the anthers and subsequent germination and development of the pollen tube to the micropyle of the ovule.
- The transfer of pollen is carried out by wind, water, and animals, mainly insects and birds.
- Wind-pollinated flowers, covered with sticky trichomes and sometimes branched stigmas, pendulous catkin inflorescences, and thin, smooth pollen grains, typically have an inconspicuous reduced perianth, long slender filaments and styles.
- Wind pollination is derived from angiosperm and has evolved in many different groups independently.
- For example, in the Heliantheae and Anthemideae tribes, wind pollination accompanied by floral reduction has independently evolved within the aster family.
- In only a few aquatic plants, water pollination occurs and is extremely complex and derived.
- There is a wide variety of angiosperm animal pollinators and a wide range of flowers adaptations to attract such pollinators.
- Beetles pollinate some of the living non-specialized families of basal angiosperm.
- The beetles feed on pieces of the perianth and stamens.
- Bees are responsible for more flowers being pollinated than any other animal community.
- Typically, bees feed on nectar and occasionally on pollen.
- By visiting flowers of several species, they may be general pollinators, or they may have modified (i.e. elongated) their mouth parts to various flower depths and become skilled in pollinating only a single species.
- Bee pollinated flowers typically have a zygomorphic, or bilaterally symmetrical, lower lip corolla that provides the bee with a landing platform.
- Either at the base of the corolla tube or in extensions of the corolla base, nectar is commonly produced.
- In orchids (e.g., Ophrys speculum), a high degree of co-evolution is common where the flower not only appears to resemble the female wasp of a specific species, but also generates the pheromone released by the insect that attract males of the species.
- By pseudo-copulation with the orchid flower, the male wasp affects pollination.
- Flies, butterflies, moths and mosquitoes are other insect pollinators.
- Since they look and smell like rotting meat, many flowers pollinated by flies are called carrion flowers.
- Birds, bats, small marsupials, and small rodents are vertebrate pollinators.
- Some bird-pollinated flowers, especially those pollinated by hummingbirds, are bright red.
- As their food source, hummingbirds depend solely on nectar.
- Bird-pollinated flowers (e.g., fuchsia) contain abundant amounts of nectar but little to no odor because birds have a very poor sense of smell.
- They normally open only at night, when the bats are the most active, and sometimes hang on long stalks of inflorescence, providing easy access to the nectar and pollen.
- Small marsupials pollinate some eucalypts (Eucalyptus)
- Whatever the agent of dispersal, when a pollen grain lands on a receptive stigma, the first stage of pollination is successful.
- The stigma surface may be wet or dry and is mostly composed of specialized glandular tissue; secretory transmitting tissue lines the style.
- Their secretions create an aura that nourishes the pollen tube as the style elongates and evolves.
- If mitosis has not yet occurred in the pollen grain in the generative cell, it does so at this point.
- Many angiosperms have developed a chemical framework of self-incompatibility to prevent self-fertilization.
- Sporophytic self-incompatibility is the most common form, where secretions of the stigmatic tissue or the transmitting tissue prevent incompatible pollen from germinating or developing.
- A second form, gametophytic self-incompatibility, includes the inability to fuse and form a zygote of the gametes from the same parent plant or, if the zygote forms, then it does not grow.
- Finally, the pollen tube passes through the micropyle via an ovule and penetrates one of the sterile cells on either side of the egg.
- Immediately after pollination, these synergids begin to degenerate.
Fertilization and embryogenesis:
- The pollen tube releases the two sperm into the embryo sac after penetrating the degenerated synergids, where one fuses with the egg and forms a zygote, and the other fuses with the central cell’s two polar nuclei and forms a nucleus of triple fusion, or endosperm.
- This is called double fertilization since another fusion process (that of a sperm with the polar nuclei) that resembles fertilization accompanies the true fertilization (fusion of a sperm with an egg).
- There is now a total chromosome complement (i.e., diploid) in the zygote and three chromosomes in the endosperm nucleus.
- To form the endosperm of the seed, which is a food-storage tissue used by the developing embryo and the subsequent germinating seed, the endosperm nucleus divides mitotically.
- It has been shown that, while they still undergo double fertilization, some of the more basal angiosperms actually form diploid endosperm.
- On the basis of when the cell wall develops, the three key types of endosperm formation found in angiosperms, nuclear, cellular, and helobial, are categorized.
- In the formation of nuclear endosperm, repeated free-nuclear divisions occurs.
- If a cell wall formation takes place, it will form after free-nuclear division.
- Cell-wall formation is associated with nuclear divisions in cellular endosperm formation.
- A cell wall is laid down between the first two nuclei in the helobial endosperm formation, during which one half forms endosperm along the cellular pattern and the other half along the nuclear pattern.
- The endosperm degenerates in many plants, however, and food is retained by the embryo (e.g. peanut, Arachis hypogaea), the remaining nucellus (e.g. beet), or even the seed coat.
- The least specialized endosperm type with nuclear and helobial forms derived from it is the cellular endosperm.
- To form a multicellular, undifferentiated embryo, the zygote undergoes a series of mitotic divisions.
- A basal stalk or suspensor forms at the micropylar end, which disappears after a very short time and has no apparent angiosperm feature.
- The embryo proper is at the end of the chalazal (the area opposite the micropyle).
- Embryo differentiation, such as the growth of cells and organs with unique functions, involves the development of a primary root apical meristem (or radicle) adjacent to the suspensor from which the root grows and the development one cotyledon (in monocotyledons) or two cotyledons (in dicotyledons) at the opposite end of the suspensor.
- A shoot apical meristem is the site of stem differentiation and differentiates between the two cotyledons or next to the single cotyledon.
- The mature embryo is a miniature plant with one or two attached cotyledons, consisting of a short axis.
- The epicotyl that extends above the cotyledon(s) includes the apex of the shoot and the primordia of the leaf; the hypocotyl, the transition region between the shoot and the root; and the radicle.
- Three different generations of angiosperm seed growth, plus a new entity are the parent sporophyte, the gametophyte, the new sporophyte, and the new one, namely, the endosperm.